Disturbance ecology
Disturbance ecology is a field of ecology that studies disturbances in the landscape. Fire is a key agent of disturbance in many parts of the world and fire ecology is a subset of disturbance ecology.
Disturbances can also be a tool for managers and sometimes there is a requirement for managers of protected areas to introduce a disturbance in order to increase or maintain biodiversity or ecological functions.

Succession
Ecological succession is the process of change in the structure of an ecological community over time. Key ideas within this concept are that nothing remains the same and habitats are constantly changing.
Primary succession is the series of community changes that occur on an entirely new habitat, which has never been colonised before. The species that colonise such habitats are termed pioneer species. An important outcome of a primary succession is to build or rebuild soils. Secondary succession is the series of community changes that take place on a previously colonized, but disturbed or damaged habitat.
Although examples of succession usually deal with plant succession, it is worth remembering that as plant communities change, so will the associated micro-organisms, fungus and animal species. The structure or the ‘architecture’ of the plant communities influences the animal species. In many ecosystems, response to disturbance is predictable as a series of stages known as seres, or seral stages. Communities change gradually and (reasonably) predictably from one sere to another.
A simplistic explanation of succession suggests that succession is directional and finally ends in a climax community. This idea suggests that succession does not progress further than the climax community, understood to be the final stage. In some ecosystems, a climax community may take hundreds of years to develop (e.g. rainforests) A more sophisticated explanation of succession recognizes that, in many ecosystems, a climax community is not the end point. In such ecosystems, disturbances are so common that a climax community never develops. For these kinds of ecosystems, although the timing of any particular disturbance may not be predictable, the fact that a disturbance will occur at some time point is almost certain.
Intermediate disturbance hypothesis
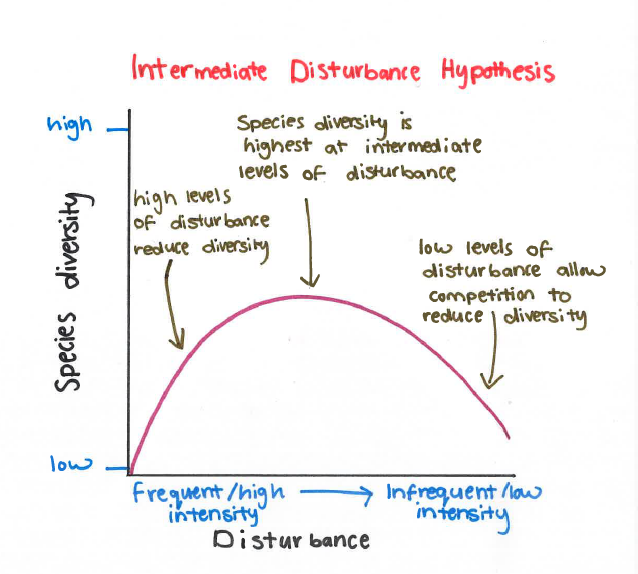
The intermediate disturbance hypothesis (IDH) states that biodiversity will be highest at intermediate levels of disturbance and lower at both high and low levels of disturbance. At very low levels of disturbance, communities are dominated by a few species which are well-adapted to the stable environment. At very high levels of disturbance, the number of species is reduced because many species cannot continue to survive the rapid changes in the environment. Intermediate levels of disturbance can provide opportunities for a wide range of species to thrive in an environment.
The IDH is sometimes criticised for its simplistic approach to the measurement of disturbance, which has many components. For example, the frequency, extent and time since the last disturbance are all interrelated measures of disturbance which could be described as ‘intermediate’, but the mechanisms underlying their effects are likely to be different.
The dynamic equilibrium model
The dynamic equilibrium model (DEM) predicts that the productivity of an ecosystem affects the relationship between disturbance and biodiversity as modeled by the IDH. Essentially the peak of the ‘hump’ of the IDH graph is shifted depending on the productivity of the ecosystem.
Describing, measuring and monitoring disturbances
While most people acknowledge two measurable components of disturbance (frequency – how often; and magnitude – how big), there are many more components. These include: seasonality (time of year), severity (extent of impact), scale (area affected) to name a few.
Winners and losers
Often a few, highly resilient species (“winners”) gradually come to dominate in disturbed environments. Other species which are not able to persist in the face of repeated disturbances do not persist in such environments. These species may be described as “losers”.
Sometimes, winners are exotic, cosmopolitan species capable of rapidly expanding their geographic range. The introduced Red Fox, Vulpes vulpes, is an excellent example of a highly adaptable species which can take advantage of disturbance events. Some native species such as Flame Robin Petroica phoenicea thrive in fire affected environments. By contrast, losers are usually represented by species with particular habitat requirements like the Greater Glider who relies on old growth trees. If their habitat is reduced or destroyed, they will not be able to adapt to the changed environment.
In some disturbance-prone natural environments, some native flora and fauna may be better adapted to a particular disturbance regime than introduced species. This is certainly seen in south-eastern Australian environments where fire is a locally common disturbance event; and native flora and fauna have adapted to its presence in the environment. In such circumstances, the local species may have a significant advantage over introduced species.
Back to the ecology theme overview







